









Рак американский сигнальный



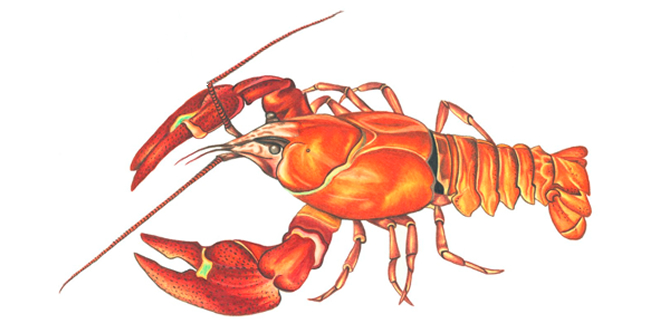
Почти все тело рака покрыто панцирем — склерозированной кутикулой, которая служит наружным скелетом. Панцирь туловища и придатков состоит из сегментов, члеников, соединенных суставной кутикулой. Обычный цвет речных раков от темно-зеленого до темно-пепельного, он варьирует в зависимости от окружающей среды. Раки, обитающие на каменистых грунтах, имеют темную окраску, на песчаных — светлую. Часто встречаются узкопалые раки нежно-голубой окраски, а широкопалые — с темно-голубой окраской внешних покровов. У голубых раков переходные участки кутикулы в суставные мембраны темного цвета. Голубые раки обитают у родников, где грунт содержит голубую глину. В литературе имеются сведения о голубых омарах и крабах. Кутикула является хорошим барьером против болезнетворных организмов, обладает анти-обрастающим механизмом. Кутикула с внутренней стороны покрыта эпителием, содержащим каратинопротеиды голубого и фиолетового цвета. При термической обработке происходит распад каратинопротеидов, кроме красного, который устойчив к термической обработке. Тело рака состоит из первичной головы (protocephalon), челестегруди, или гнатоторакса (gnathothorax), груди (thorax), брюшка (abdomen) и тельсона (telson). Протоцефолон, гнатоторакс и торакс покрыты общим щитом — карапаксом (carapax). Передняя часть тела заканчивается рострумом (rostrum), по бокам которого находятся глазные вырезки (incisurae oculi). На панцире видны три шва: затылочный (suturae cervicalis), отделяющий головную часть от груди, и два жаберно-сердечных (suturae branchiocardiale). Боковые части панциря — бранхиостегиты (branchiostegit) образуют жаберные полости. Брюшко состоит из шести подвижно сочлененных сегментов и тельсона. Каждый сегмент с дорсальной стороны покрыт выпуклой пластинкой-тергитом (tergum), а с вентральной ограничен стернитами (sternum). Речной рак имеет 19 пар конечностей (5 головных, 8 грудных и 6 брюшных). Характерные головные придатки — антенны и двуветвистые антеннулы, а также верхние и нижние челюсти(mandibulae et maxillae), — преоральные по своему происхождению. Грудными придатками служат три пары ногочелюстей (maxillepedes) и пять пар ходильных конечностей (pereiopodae). Все придатки имеют специализированное назначение. Они обеспечивают питание, добычу пищи, защиту, дыхание, ориентацию, вынашивание потомства и др. Большой интерес представляют для исследователей строение и функция конечностей речного рака как прообраз манипулятора подводного робота. Первые три пары ходильных ног снабжены клешнями и называются хелипедами (chelipedae), две последние пары заканчиваются коготками. Конечности, снабженные клешнями, выполняют важные функции в жизнедеятельности речного рака, а именно в добыче пищи, питании, обороне и др. Все конечности имеют общую схему строения для ракообразных и состоят из четырех отделов протоподита (protopodit), экзоподита (exopodit), эндоподита (endopodit) и эпиподита (epipodit). От коксоподита (coxopodit) отходит эпиподит. Базоподит сочленен с экзоподитом и коксоподитом. Эндрподит содержит 5 члеников: базоподит (basipodit), ишиоподит (ischiopodit), мероподит (meropodit), карпоподит (carpopodit), проподит (propodit), дактилоподит (dactylopodit). Ходильных ног (pereiopodae) у речного рака 5 пар, непосредственно в передвижении участвуют 4 пары конечностей. Первая пара является вспомогательной в преодолении препятствий. Периоподы лишены экзоподитов и имеют одноветвистое строение. На эпиподитах периоподов имеются по одной ветви жабр и пучки нитевидных выростов. Брюшными ножками (pleopodae) снабжены первые пять сегментов, шестой сегмент представлен плоскими пластинами — уроподами. Первая пара ножек у самцов превращена в капулятивный орган — гоноподы. Вторая пара брюшных ножек также участвует в выводе сперматофор, и поэтому их иногда называют вторыми гоноподами. У самок первая пара брюшных ножек лишена экзоподита. Остальные ножки имеют двуветвистое строение. Придатки шестого сегмента — уроподы — состоят из протоподита, экзоподита и эндоподита.
Жизненный цикл сигнального рака сходен с таковым у рассмотренных выше видов. Спаривание происходит в октябре, однако, в отличие от других видов, не зависит от температуры (Abrahamsson, 1971). Сроки вылупления молоди связаны с температурой: в реках и ручьях штатов Калифорния, Орегон и Вашингтон выклев молоди происходит в апреле — мае, а в холодноводном оз. Тахо — в июле — августе (McGriff,1983). Продолжительность созревания молоди сильно варьирует и, вероятно, тоже зависит от температуры: в Дип-ривер (штат Вашингтон) — в течение 1 года, в устье р. Сакраменто (штат Калифорния), в шведских прудах, в водоемах Польши −1-2 года, в оз. Тахо (Калифорния) и р. Берри-крик (штат Орегон) — 2-3 года (McGriff,1983). По общему мнению исследователей, сигнальный рак характеризуется более высокими показателями роста, чем широкопалый (Abrahamsson, Cukersis, 1978): в водоемах Литвы, например, сигнальный рак достигает промысловых размеров за 2 года, тогда как широкопалый — за 4 года. Плодовитость сигнального рака (длиной 9-10 см) находится примерно на том же уровне, что и у широкопалого. Сигнальный рак по продукционным характеристикам представляет собой несколько более конкурентоспособную форму, нежели широкопалый. При вселении сигнального рака в водоемы Швеции (55-600 с.ш.) широкопалый рак из этих водоемов, как правило, исчезал (Furst, 1984). Выпадение аборигенного вида происходило, по-видимому, вследствие вспышки «рачьей чумы» (Abrahamsson, 1971; Цукерзис, 1977). Несколько иная картина наблюдалась в водоемах Финляндии (60-630 с.ш.), куда тоже был вселен сигнальный рак. Исследователи отмечают более медленный рост численности P. leniusculus, чем в водоемах Швеции (Westman, Pursiainen, 1984). В случае сосуществования сигнального и широкопалого раков в одном водоеме численность первого стабилизировалась на очень низком уровне — в 7 раз ниже, чем у аборигенного вида (Westman, Pursiainen, 1978). Можно предположить, что на северной границе ареала конкурентное преимущество сигнального рака ослабевает. В Финляндии, куда производилось вселение сигнального рака, и Ленинградская область находятся в пределах одних широт (60-630 с.ш. и 59-620 с.ш. соответственно).
Северо-американский вид. Благодаря высоким вкусовым качествам и значительной доле съедобных частей тела в общей массе особи этот вид высоко ценится на международном рынке и служит объектом акклиматизационных работ в странах Европы.
Географические границы ареала данного вида точно не установлены. Популяции сигнального рака, изучаемые американскими исследователями, находятся на западном побережье США, в пределах штатов Калифорния, Орегон, Вашингтон — между 30 и 50о с.ш. (Abrahamsson, Goldman, 1970; McGriff, 1983)/ Мамо (Mamot. 1984) относит данный вид к среднеширотным, а рассмотренные выше виды речных раков (широкопалый и длиннопалый) — к высокоширотным. При акклиматизации сигнального рака на Северо-западе Европы (Швеция, Финляндия) северная граница его распространения проходила по 63о c.ш. (Westman, Pursiainen, 1978/ 1984; Furst, 1984). В Северной Америке сигнальный рак обитает в основном в реках и ручьях. В начале века данный вид был вселен в глубоководное оз. Тахо, где успешно прижился и освоил каменистую «литораль» вплоть до глубин 40 м (Abrahamsson ,Goldman, 1970; McGriff, 1983). В Западной Европе вселение этого вида проводили, как правило, в непроточные водоемы, ввиду того, что он является переносчиком губительной для аборигенных видов «рачьей чумы». По общему мнению исследователей, необходимым условием для обитания данного вида в водоеме служит наличие каменистого грунта (Westman, Pursiainen, 1978; Flint et al., 1977; Daniels, 1980). Из трех рассматриваемых видов речных раков, сигнальный, вероятно, является самым стенобионтным в отношении характера грунта; он встречается только на каменистой литорали даже в условиях конкурентных взаимоотношений с широкопалым раком (Westman, Pursiainen, 1978). По данным экспериментальных работ Масона (Mason, 1978) такая приверженность сигнального рака к каменистому грунту объясняется необходимостью укрытий, вследствие характерного для данного вида каннибализма. Данные об отношении рассматриваемого вида к гидрохимическим факторам практически отсутствуют: американские исследователи этому вопросу не уделяют внимания, а европейские проводят акклиматизационные работы с этим видом исключительно на олигосапробных водоемах, где, по их мнению, мог бы обитать аборигенный широкопалый рак. По отношению к минерализации воды сигнального рака можно отнести к эвригалинным формам: он может существовать в морской воде (до 250/00) и хорошо приживается в маломинерализованных (менее 0,10/00, 100 мг/л) озерах Финляндии (Westman, Pursiainen, 1978;Randquist, Goldman, 1978). Данных об отношении сигнального рака к температуре тоже мало. Зона оптимальных температур (для питания, а, следовательно, и для роста) лежит в диапазоне от 20 до 280 С (Strempel.1976) и сходна с таковой у A. leptodactylus. Выклев молоди P.leniusculus происходит при температуре 18-200С (Abrahamsson ,Goldman, 1970). Следует отметить, что северная граница расселения сигнального рака в Финляндии проходит, примерно, по той же широте (630 с.ш.), что и северная граница ареала A. leptodactylus (620 с.ш.), что вероятно, связано со сходным отношением рассматриваемых видов к температуре.